时间:2018-08-13 点击: 次 来源:易邦生物 作者:刘东,宋姗姗等 - 小 + 大
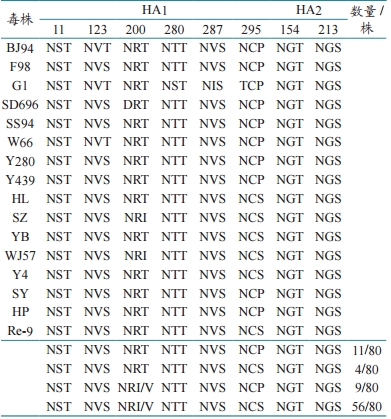
4 个毒株(4/80)含有 8 个潜在的糖基化位点,而且这 4 个毒株只有在 2013 年和 2014 年分离得到(表 3),9 个毒株含有 6 个糖基化位点,也是2013 年和 2014 年的分离株。与 Y280 相比,其他毒株在第 203 位氨基酸大多发生了 T → I/V,导致了第 200 位糖基化位点逐渐缺失,第 297 位氨基酸发生了 P → S 的变化,使得 295 位多出了一个潜在的糖基化位点 NCS。新的糖基化位点的出现或者原有糖基化位点的缺失均有可能通过改变受体结合位点和 HA 蛋白的抗原性来感染新宿主。 表 3 H9 亚型 AIV HA 蛋白糖基化位点 3 讨论 H9N2 亚型 AIV 在世界范围内广泛分布,在遗传进化上,可以分为北美系和欧亚系两大分支,而欧亚系又进一步衍生出以 BJ/94-like 或 Y280-like、G1-like、Y439-like 以及 F/98-like 等为代表的病毒亚群。根据新的亚系命名系统,我国 H9N2 亚型AIV 分离毒株主要来源于 H9.4.2 分支,而 4.2 分支又演化为 4.2.1~4.2.6 分支。本次分离到的 80 株H9N2 亚型 AIV 毒株均属于 H9.4.2.5 分支的独立(Y4)亚群。 本研究分析了 2013—2018 年分离得到的 80株 H9 亚型AIV 的 HA 基因序列, 发现这80 株HA 基因的裂解位点序列大多为 PSRSSR,与参考毒株 Y280 不同,其第 2 位氨基酸残基由非极性氨基酸(A)突变为极性氨基酸(S)。氨基酸序列分析发现,这 80 株病毒在 HA 裂解位点序列的第5 位氨基酸残基均为 S,由 AGT 编码,如果发生A 替换 T/C,该位点的 S 就会变为 R,那么这些毒株就会变为高致性病毒株,因而具有潜在毒力增强的风险。 毒株的潜在糖基化位点也是影响毒株毒力的关键位点之一。在本研究分离的 80 株毒株中,2013—2014 年有 4 株含有 8 个潜在糖基化位点,与 F98、Y280 等参考毒株相比多出了 295NCS 这一潜在糖基化位点,有 9 株含有 6 个潜在糖基化位点,与 F98、Y280 等参考毒株相比消失的潜在糖基化位点是 200NRT,而其余分离株,特别是2015 年以后的毒株,在第 203 位氨基酸大多发生了 T → I/V 的变化,导致了第 200 位糖基化位点逐渐缺失,第 297 位氨基酸发生了 P → S 的变化,使得 295 位多出了一个潜在的糖基化位点 NCS。新的糖基化位点的出现或者原有糖基化位点的缺失均有可能通过改变受体结合位点和 HA 蛋白的抗原性来感染新宿主。 目前灭活疫苗免疫仍是防控 H9N2 亚型禽流感的主要方法。该方法虽然起到了一定的防控作用,但由于疫苗的长期使用,导致免疫压力持续存在,使得 H9N2 亚型 AIV 可能会不断重组和变异。因此,疫苗株的选择一定要与现场流行毒株匹配,否则就起不到理想的保护作用。为及时了解和掌握H9N2 亚型 AIV 的流行动态及变异情况,必须做好病毒的长期跟踪监测工作,这样可以根据病毒的流行和变异情况,及时研发有效的针对性疫苗,确保H9 亚型 AI 的防控效果。 |
上一篇:承德市接坝地区包虫病流行病学调查
|
建议使用1440*900分辨率浏览