时间:2021-04-03 点击: 次 来源:广东省畜牧兽医学会 作者:金宣讲 - 小 + 大
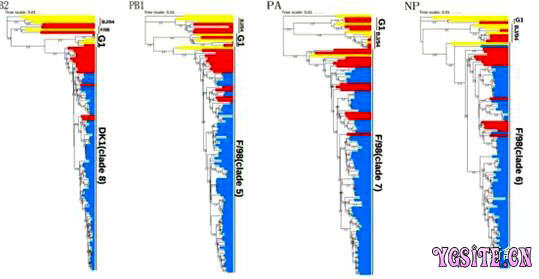
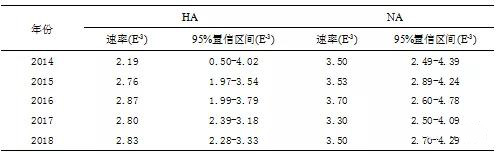
2. 内部基因的系统进化 H9N2内部基因的进化支先前由宿主来源(forexample, Dk1, Dk2, Dk3, Aq, Swine and unknown avian lineages),代表性菌株(forexample, BJ/94, G1, F/98 and Ty/ 66 lineages),地理位置(Koreanlineage)或病毒亚型(forexample, H5N1-Gx/Gd, H5N1-Z and H5N1-like lineages)命名。在这里,我们使用数字根据内部基因的系统发育命名每个进化支。所有六个内部区段被分为42个独立的进化支。 本研究中M基因全部由G1-like病毒进化而来,被分配在clade 3(图1)。我们观察到所有病毒的NS基因全部由A/chicken/Beijing/1/94病毒进化而来,被分配在clade 7(图1)。PB2基因都是从Dk1-original谱系进化而来,被分配到clade 8(图2)。所有毒株的PB1,PA,NP基因都是从SF/98-like谱系进化而来,在各自的进化树中他们又分别被分配到clade 5,clade 7,clade 6(图2)。与外部基因相比,所有病毒的六个内部基因在各自的进化树中都分别属于同一个分支。病毒6个内部基因的分支组合都能够稳定遗传,说明这种内部基因的组合方式适应性强,更利于病毒本身在自然中生存。 图2 广东省H9N2亚型禽流感病毒PB2/PB1/PA/NP基因遗传进化树 3. 基因型 根据72株病毒8个片段系统发育分析,划分他们的基因型。我们一共发现了3种基因型(G57,G68,G118),其中新发现的一种被命名为G118(图3)。基因型G57是我们研究中的主要基因型,其余2种基因型仅在2015-2016期间偶尔在广东检测到。与G57基因型相比,G68基因型仅HA基因所在的分支不同,新出现的G118基因型仅NA基因所在的分支不同。这很有可能是两种不同基因型毒株感染同一宿主时交换NA基因,进行了重配。尽管新型基因型(G118)仅偶尔在家禽中检测到,但我们的研究中基因型的数量有所增加,这表明2014- 2018年广东的H9N2禽流感病毒变得越来越多样化。 4. 进化速率 我们计算了2014-2018年H9N2流感病毒HA和NA基因的核苷酸替代率。可以看出,HA基因和NA基因各年份核苷酸替代率的范围分别为2.19E-3-2.87E-3和3.30E-3-3.70E-3。值得注意的是,HA基因核苷酸替代率在2015,2016年异常高。由遗传进化树分析结果可知2015,2016年的H9N2病毒中分别有一株毒株的HA基因与其他毒株不在同一分支。同样值得注意的是NA基因核苷酸替代率仅在2015异常高。而2015年的H9N2病毒中有8株毒株的NA基因与其他毒株遗传距离较远。 表1 G57基因型两个时期的核酸替换速率 |
上一篇:非洲猪瘟的检测方法分析
|
建议使用1440*900分辨率浏览