时间:2018-08-13 点击: 次 来源:易邦生物 作者:刘东,宋姗姗等 - 小 + 大
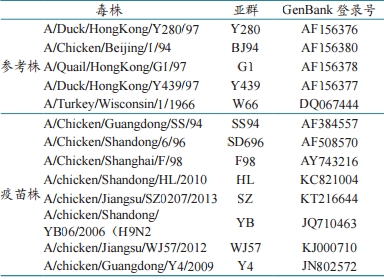
2.2 HA 基因遗传进化分析 从 GenBank 中选取 5 个 H9 亚型代表毒株及11个我国普遍使用疫苗毒株的 HA 基因序列(表1),与分离的 80 株毒株 HA 基因进行对比和遗传进化树分析(图 1)。从图中可看出 2013—2018 年分离到的 H9 亚型 AIV 大部分在 H9.4.2.5 分支上,只有2013 年云南的 A-CK-yunnan-2013 在 H9.4.2.6 分支上。H9.4.2.5 分支又有许多小分支,其中:79 个分离株在 Y4 亚群,与 Y4 疫苗株的核苷酸同源性在93.7% 以内;60 个分离株在 WJ57 亚群,与 WJ57疫苗株核苷酸同源性在 95.5% 以内。遗传进化树分析显示:2013—2018 年的分离毒株在遗传进化树上基本按时间顺序排列,而且随着时间推移,新分离毒株逐渐远离疫苗毒株。 表1 选用的 H9 亚型 AIV 参考株 2.3 HA 基因关键氨基酸变异分析 参照人源 H3N2 流感病毒 A/Achi/2/1968 毒株的 HA 基因氨基酸序列,推导并比较 80 个毒株HA 基因关键位点的氨基酸序列。结果显示:80 株病毒 HA 基因裂解位点氨基酸序列(321~329)中,除 1 株为 PSRSSN、1 株为 PSRSNR 外,其余 78株均为 PSRSSR,均符合低致病性禽流感特征。这些毒株的裂解位点氨基酸序列虽为低致病性特征,但不同于参考毒株 Y280(PARSSR),其第 2 位氨基酸残基由非极性氨基酸(A)突变为极性氨基酸(S)。通过核苷酸序列分析发现,这 80 株毒株 HA 裂解位点序列的第 5 位氨基酸残基均为 S,由 AGT 编码。 H9 亚 型 AIV HA 基因在 109、161、163、191、198、202、203 和 146~150 及 232~237 位(H9序列位置)构成受体结合位点的袋状结构。80 株病毒基因受体结合位点右侧壁(146~150)比较保守,主要模式为 GTSKA,受体结合位点左侧壁(232~237)中的 234 和 235 位正在发生突变,主要模式为 NGLMGR,而国内参考毒株 Y280 的模式为 NGLQGR,BJ94、F98 的模式为 NGQQGR。受体结合位点的改变主要是在 198 位,呈现 T/V/A/K 的突变,226 位氨基酸位点与唾液酸受体α-2,6Gal 和 α-2,3Gal 的结合特性有关。本研究中的所有 H9N2 亚型 AIV 的 226 位氨基酸均为亮氨酸(L),而早期的大多为谷氨酰胺(Q)。这说明2013 年以后的 H9N2 病毒具备了结合 α-2,6Gal 唾液酸受体的能力,病毒感染哺乳动物,甚至是人的概率大大增加(表 2)。 表 2 H9 亚型 AIV 裂解位点和袋状结合区的突变 |
上一篇:承德市接坝地区包虫病流行病学调查
|
建议使用1440*900分辨率浏览