时间:2022-02-17 点击: 次 来源:饲料工业杂志 作者:张佳,王园 - 小 + 大
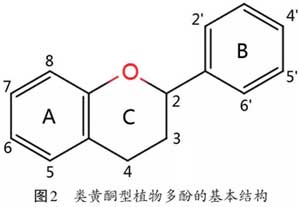
2、PP的生物活性及其作用机制 2.1 抗氧化活性 抗氧化活性是PP研究最为深入的特性。体外研究证实,PP对DPPH、羟基自由基、超氧阴离子自由基具有很强的清除活性。目前研究发现,PP中抗氧化活性最高的物质是杨梅酮。Barzegar(2016)以二氯二氢荧光素(DCF)探针作为MCF-7细胞内活性氧(ROS)的指示剂,发现2 μg和5 μg的杨梅酮均可直接抑制细胞内ROS的产生。这与PP的结构密切相关,如图2所示:①B环上的邻苯二酚基团具有显著的清除ROS或活性氮(RNS)的能力;②B环上的羟基是自由基的电子和氮的供体;③C环可以使苯氧自由基更具稳定性,增强抗氧化活性;④当A环和C环上存在4-OH时,7-OH具有最大的自由基清除能力;⑤A环的5-OH和7-OH基团可以与金属离子络合,增加自由基清除能力。目前PP螯合金属离子,抑制氧化应激产生的能力已得到证实。Bong-Jeun(2004)研究发现,绿茶多酚可以螯合金属离子(Cu2+、Fe2+),加速阻止O2-和H2O2变成羟自由基(·OH)。当其浓度在5 mg/kg以上时,Cu2+的螯合效果可达80%以上,而在50 mg/kg以上时,Fe2+的螯合效果可达70%左右。不仅如此,近年来国内外学者通过对PP抗氧化机制的深入研究,发现PP还可以通过核因子E2相关因子2(Nuclear factor-erythroid 2-related factor-2,Nrf2)-抗氧化反应原件(antioxidant responsive element,ARE)信号通路诱导抗氧化酶或Ⅱ相解毒酶表达上调。买地娜依等(2018)研究发现,1、10、50 μg/mL的表没食子儿茶素没食子酸酯(EGCG)均可以上调3T3-L1脂肪细胞Nrf2和HO-1的基因表达量,进而提高细胞内谷胱甘肽(GSH)、超氧化物歧化酶(SOD)含量,降低丙二醛(MDA)含量,抑制细胞发生氧化应激,且呈剂量依赖性。进一步研究表明,EGCG可激活丝裂原激活蛋白激酶(mitogen-activated protein kinases,MAPK),触发Nrf2磷酸化,或直接与Kelch样环氧氯丙烷相关蛋白1(Kelch-like ECH-associated protein 1,Keap1)中的半胱氨酸残基互作,从而诱导Nrf2解离。此外,Gan等(2016)证实25 μmol/L白藜芦醇可通过ERK5/HDAC5途径激活Mn-SOD的表达,从而减轻内皮细胞线粒体氧化应激。Liu等(2018)发现,蓝靛果多酚(0.05%和0.1%)可有效促进高脂饲粮诱导的小鼠肝脏内Nrf2、HO-1和MnSOD等抗氧化蛋白的表达,从而降低脂质过氧化水平。综上所述,PP参与调控抗氧化作用的调节机制主要包括:①清除自由基;②螯合金属离子抑制活性氧的形成;③激活Nrf2-ARE、ERK5/HDAC5等信号通路,诱导抗氧化酶或Ⅱ相解毒酶表达。 2.2 抗炎活性 体内、外试验均已证明PP可通过抑制促炎细胞因子的表达实现其抗炎活性。蓝莓花色苷、黑莓花色苷和黑醋栗花色苷(20 μg/mL)能显著降低脂多糖(LPS)诱导的小鼠巨噬细胞白介素-1β(IL-1β)、肿瘤坏死因子α(TNFα) mRNA的表达水平及含量。绿茶多酚(1 mg/mL)则可以抑制Caco-2细胞(人结直肠腺癌细胞)、白细胞介素-6(IL-6)和白细胞介素-8(IL-8)的释放,降低细胞通透性,预防炎症性肠道疾病。在炎症过程中,诱导型一氧化氮合酶(iNOS)和环氧化酶2(cyclooxygenase,COX-2)及各自催化产物一氧化氮(NO)和前列腺素E2(PGE2),参与并加重炎症。Koeberle(2009)发现姜黄素(30 μmol/L)可以通过抑制COX-2的表达和直接激活mPGE2活性来抑制PGE2的产生。蔡美云等(2020)进一步发现,80 μmol/L的山奈酚(KAE)可通过抑制P38 MAPK/NF-κB信号通路,显著降低iNOS和COX-2的表达,进而减轻神经毒素6-羟多巴胺诱导的PC12细胞炎症反应的发生。NF-κB原本与I-κBα抑制剂结合存在于胞浆中,但当细胞受到炎症因子刺激时,I-κBα被磷酸化和降解,从而释放NF-κB进入细胞核,诱导促炎基因表达。研究发现,PP可以抑制NF-κB核转位,如从黑豆中分离出的花青素就可以有效抑制NF-κB转位到细胞核中。故PP发挥抗炎活性的途径有:①抑制促炎细胞因子的表达;②抑制MAPKs及NF-κB信号通路介导iNOS和COX-2的表达,进而抑制炎症;③抑制NF-κB核转位。 |
上一篇:牧草引种、育种与丰产栽培技术
下一篇:饲料生产过程中的水分控制
|
建议使用1440*900分辨率浏览