时间:2018-06-08 点击: 次 来源:中国动物检疫 作者:董李学,袁万哲,左玉柱 - 小 + 大
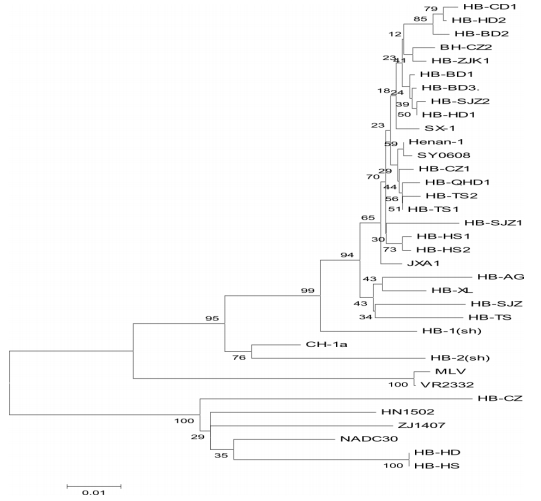
图3 ORF5 基因遗传进化树 2.4 ORF5 基因毒力和主要抗原表位变异分析 ORF5 第 151 位氨基酸位于糖蛋白的疏水区。此氨基酸的变异改变了该区域的疏水性,从而可能改变病毒毒力。研究结果显示,第 13 和 151 处的氨基酸突变与毒株毒力有关。与北美型标准株 VR-2332 相比,本试验获得的 23 个毒株中,HB-HS、HB-HD、HB-CZ 的第 13 位均发生变异(突变为P 或 Q),其他未发生变化,均为 R;HB-SJZ1、HB-HD、HB-CZ、HB-HS、HB-HS1、HB-HS2 的第 151 位已突变为赖氨酸(K)。这可能导致了该毒株毒力发生改变。而 HB-HS、HB-HD、HB-CZ同时存在第 13 和 151 处的氨基酸突变。对 ORF5基因的 RT-PCR 产物,用内切酶 Mlu Ⅰ进行 RFLP分析,以区分疫苗株和其他北美毒株。这是由于疫苗株第 137 位氨基酸为 Ala,而其他野毒株的第137 位均为 Ser。本试验获得的 23 个 ORF5 氨基酸序列中的第 137 位氨基酸均为 Ser,表明所获得的 23 个毒株均可能为野毒株。目前已确定的美洲株表位有 3 个:2 个为非中和表位(27~30和180~197),1个为中和表位(37~45)。 本试验获得的 23 个 ORF5 氨基酸序列分析表明,在非中和表位 29 位点处,除 HBBD3、HB-SJZ、HB-SJZ1、HB-HD、HB-CZ、HB-HS、HB-HS1、HB-HS2、HB-XL、HB-AG 为 Val 外, 其余均为Ala(与经典株 VR-2332 和疫苗株 MLV 相同);在 180~197 位点中,第 185 位点均为 Ala,与国内分离的 HP-PRRSV 毒 株 SY0608、JXA1、SX-1、Henan-1 相 同,VR-2332、MLV、CH-1a 株均为Val; 第 189 位点分别均为 Leu 或 Val, 而 VR-2332、MLV 株则为 Ile;HB-BD2 第 194 位点突变为 Gly;HB-SJZ1 第 191 位点突变为 Ile,HB-HS、HB-CZ、HB-HD 为 K。在中和表位(37~45)中,除 HB-HS、HB-CZ、HB-HD 为 Leu 外, 第 39 位点均为 Ile,VR-2332、MLV 株为 Leu,而 CH-1a、HB-2(sh)株则为 Phe,NADC30 株为 Leu。这说明 23 个流行毒株的氨基酸序列中抗原表位已发生变异,其中以 HB-HS、HB-CZ、HB-HD 株变异最大。 3 讨论 自 2006 年发生 HP-PRRS 以来,河北省大中型猪场不断采取以疫苗免疫为主的积极预防措施。但鉴于 PRRSV 的高变异及重组,需要不断研究病毒的遗传变异趋势,从而保证疫苗防疫的有效性。从本次调查结果看,河北省 23 个流行毒株与国内流行的高致病性毒株缺失情况相同,均在 481 位和532~560 位缺失 30 个氨基酸。它们之间的同源性为89.5%~98.8%。调查中未发现经典毒株,原因可能与本研究所选择的发病猪场有关。因此,可以认为目前在河北省主要以 NSP2 基因组氨基酸缺失且位置相同为特征的美洲型 PRRSV 毒株流行为主。 |
下一篇:猪场伪狂犬病净化成效及体会
|
建议使用1440*900分辨率浏览