
禽流感、肠道菌群和免疫之间的关系(图)
|
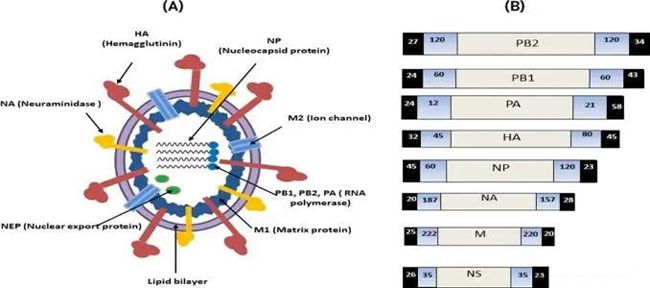
图 1:(A) IAV 的病毒体结构和 (B) 基因组组织AIV 的分类、形态和组成 AIV 属于正粘病毒科,它们是单链八节负义 RNA ( -ssRNA ) 病毒,编码至少十种病毒蛋白。该家族分为 A、B 和 C 型流感的三个主要属和另外两个未知属,isisavirus和thogotovirus 。在其他流感病毒中,属于甲型流感病毒属的 AIV 传播最为广泛,其成员感染了禽类和哺乳动物物种。AIV 在形态上是可变的,这种可变性由基质蛋白 M1 控制。 病毒是多形的,大致呈丝状,长度为几微米或球形,大小为 80 至 120 nm(图 1)。任何形态形式的病毒粒子都被来自宿主细胞的脂质包膜包裹着,具有球状刺突或膜蛋白突起(图 1)。尽管基因组大小很小,但 AIV 已经进化出各种分子方法来表达来自单个基因片段的几种病毒蛋白。 AIV 的八个 RNA 片段应该编码十种蛋白质(Kang 等人,2021 年)。 甲型流感病毒根据其包膜蛋白、神经氨酸酶 ( NA ) 和血凝素( HA )进一步根据毒株或亚型进行分类。 目前,已知 NA 和 HA 分别有 9 种 (N1-N9) 和 16 种 (H1-H16) 亚型 。HA 和 NA 都可以引发亚型特异性免疫反应,防止由相同亚型引起的感染,并部分防止由不同亚型引起的感染 。 AIV的肠嗜性 禽流感是一种世界范围的人畜共患病,通常伴随着家禽业的严重破坏以及流行病和大流行。众所周知,低致病性禽流感(LPAI)的家禽肠嗜性高于高致病性禽流感(HPAI),而高致病性禽流感(HPAI)与呼吸道上皮细胞的联系更密切。然而,由 HPAI H5N1 引起的感染对迁徙的大天鹅的肠道微生物群或粪便细菌群落的影响导致了 GIT微生物群的破坏结构由乳酸杆菌和气单胞菌等细菌属的优势修饰介导。粪便微生物组的这种组成和特征变化可能是由于肠道相关感染加剧疾病传播的有害影响。 此外,高致病性禽流感 2016 H5N8 病毒对野鸭肠道上皮细胞的附着程度与低致病性禽流感 H4N5 病毒相当。2016 年 H5N8 可能获得了与低致病性禽流感 LPAI 病毒平行的肠嗜性,而不会失去鹅/广东谱系的旧高致病性禽流感 HPAI 病毒的嗜肺性。 2016 H5N8 的肠嗜性升高意味着该病毒在野生水禽宿主中存在较长时间(Caliendo 等人,2020 年)。 此外,H9N2 是对公共健康的重大威胁,因为它可能在以前没有驯化过的哺乳动物细胞中繁殖,并在人类中繁殖 HPAI 病毒被证明具有来自禽 H9N2 病毒的内源基因 。LPAI H9N2 亚型已达到人畜共患比例,导致火鸡、鸡、家鸭、鸵鸟和野鸡感染。因此,为了提高禽类的生产力和健康,减少和控制 H9N2 爆发可以在减少哺乳动物接触病毒方面具有关键优势。 |
上一篇:全球有机产品认证体系盘点(图)
下一篇:冬季鸡传染性鼻炎的防治(图)